

В данной статье дан сравнительный анализ эмбрионального развития зубов человека и мыши.
Использованы основные критические периоды развития зубов, характерные для млекопитающих. Работа выполнена для решения вопроса о возможности использования стволовых клеток для выращивания с помощью клеточных технологий утраченных зубов.
Установлено, что развитие зубов человека по срокам дифференцировки клеток, специализации и сигнальным межклеточным взаимодействиям имеет отличия.
Ключевые слова: зубной мешочек, пульпа, одонтобласт, стволовые клетки, зубная пластинка, почка зуба, сигнальные молекулы, дифференцировка, цементобласт.
Список сокращений:
FGF- фактор роста фибробластов
BMP — костный морфогенетический белок
Eda — эктодисплазин
Hedgehog — название сигнального пути
Shh — sonic hedgehog, ген, кодирующий белок семейства hedgehog
TGFβ — трансформирующий фактор роста β
WNT — белок сигнального пути
Введение
На сегодняшний день перед учеными стоит вопрос об использовании стволовых клеток для регенерации зубов человека, так как потеря зубов остается одной изсамых существенных проблем в стоматологии. Поданным ВОЗ, полная потеря зубов вособенности широко распространена среди пожилых людей. Вглобальных масштабах примерно у50% людей ввозрасте 60 лет и старше отсутствуют зубы попричине воспалительных заболеваний пародонта ипатологии твердых тканей зубов. Неисчерпаемым источником стволовых клеток для выращивания зубов могли бы служить экспериментальные лабораторные животные, у которых регенерация зубов продолжается и в постнатальный период онтогенеза. Учёные гипотетически рассматривают использование стволовых клеток пульпы грызунов, но существующий ряд различий в строении предполагает углубленные исследования в этом направлении. Поиск сигнальных молекул, индуцирующих непрерывную физиологическую регенерацию зубов и их активация у человека создает перспективы успешного решения проблем адентии.
Материалы и методы исследования
Проведён анализ литературных данных, представленных в зарубежных и отечественных базах данных. Рассматривались вопросы изменения сигнальных взаимодействий в дифференцирующихся клетках формирующихся твердых и мягких тканей зубов на
этапах закладки, гистогенеза и специализации. Изучена молекулярная генетическая составляющая изменений генома в клеточных ансамблях развивающегося зуба. Из общего количества источников были определены 7 публикаций, как наиболее значимые. Использован архив гистологического материала эмбриона человека научного руководителя Рева Г.В.
Результаты исследования
Анализ зарубежных исследований показал, что основные результаты были получены на лабораторных животных, а не на человеке. При изучении гистологического развития зубов, на определенных этапах формирования были выявлены ряд сходств и различий:
1)Утолщение эпителия
При изучении эмбрионов человека и мышей выделены схожие эпителиальные утолщения на еще не сросшихся лицевых отростках.(рис. 1,2) Утолщения срастаются в результате слияния лицевых отростков. Различием является, что верхний боковой резец человека имеет двойное происхождение, поскольку в него входят два лицевых отростка (медиальный носовой и верхнечелюстной).[1] У мышей утолщенный эпителий покрывает всю ротовую поверхность верхней челюсти на ранних стадиях и дает начало не только зубной пластинке, но и небным гребням и преддверию рта.


Рис.1 Эмбрион человека в конце 3-й недели. Окраска гематоксилином. Увеличение х100.
Стадия а) 10; б) 30 сомитов. 2-х слойная эктодерма туловищного отдела зародыша человека (указана зелеными стрелками), вдается между сомитами более чем на 1/3 зародыша. Голубая, красная и зеленая звёздочки соответственно указывают дерматом, миотом и склеротом. Красные стрелки указывают на энтодерму. а) мезодерма в начале дифференцировки на дерматом, миотом и склеротом; б) выраженная дифференцировка мезодермы на дерматом, миотом и склеротом. Микрофото.
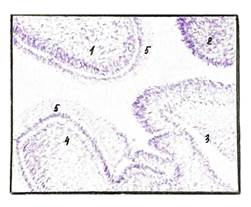
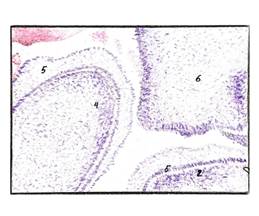
Рис. 2 Полость рта эмбриона человека пяти недель. Окраска гематоксилином и эозином. Ув. х200
1,2) носовой и медиальный небный отростки формирующейся верхней челюсти; 3) язык; 4) нижняя челюсть, 5) многослойный плоский неороговевающий эпителий.
2)Зубная пластинка
При формировании зубной пластинки утолщенный эпителий инвагинирует в соседнюю мезенхиму, что приводит к накоплению множества сигнальных сетей.[2] Во время встраивания слоя зубного эпителия в мезенхиму поверхностные эпителиальные клетки мигрируют, заполняя формирующуюся канавку. Локальные вариации физических или механических раздражителей из тканевого микроокружения также могут быть вовлечены в происхождение зубной пластинки.
Зубная пластинка представлена U-образным эпителиальным гребенем в зародышевых дугах верхней или нижней челюстей, параллельно и снаружи от нее проходит вестибулярная пластинка, которая является источником преддверия полости рта. На ранних стадиях одонтогенеза у эмбрионов отсутствуют непрерывные пластинки. Эпителий этих пластинок имеет общее происхождение в области губ нижней челюсти, но верхней челюсти приведённый ранее эпителий возникает отдельно. У мышей в резцовой и щечной области происходит раздельное загибание эпителия, то есть отсутствие непрерывной зубной пластинки.
3)Почка зуба
Почка зуба имеет крупные клетки на базальной мембране и более мелкие в центре. Одна из его медиальной и латеральной мезенхимальных граней имеет выступ. Зачатки зубного ряда начинаются как припухлости на холмике зубного эпителия. Эпителиальная почка инвагинирует на своем кончике, и ее цервикальные петли охватывают мезенхиму зубного сосочка, которая дает начало пульпе зуба и одонтобластам.[3] У эмбрионов человека и мыши в области губ верхней и нижней челюсти: параллельно зубным зачаткам появляются выпуклости вестибулярного эпителия, что указывает на взаимосвязь между сегментацией вестибулярным эпителием.
4) Стадия колпачка
На данной стадии продолжается конденсация вокруг почки и происходит деление на два клеточных пула: центральный сосочек, образующийся из нервного гребня и периферический фолликул-из мезодермы. Первый окружается зубным эпителием и даёт начало пульпе зуба и одонтобластам, а второй даёт начало цементобластам, остеобластам и фибробластам. Цементобласты формируют цемент зуба, остеобласты — альвеолярную кость, а фибробласты — периодонтальную связку. Во время стадии колпачка начинает формироваться коронка зуба. Данный процесс регулируют первичные и вторичные эмалевые узлы. Форма коронки фиксируется благодаря тому, что на эпителиально-мезенхимальной границе эпителиальные клетки дифференцируются в амелобласты, а мезенхимальные — в одонтобласты. Амелобласты секретируют продукты эмали, а одонтобласты — дентина.[4] На стадиях колпачка и колокола эпителий уже начинает окружать подлежащую мезенхиму и формировать так называемые цервикальные петли. Слой базальных эпителиальных клеток петли, ограничивающий зубной сосочек, назвали внутренним эмалевым эпителием, который дифференцируется в амелобласты. Другая часть, контактирующая с зубным фолликулом, наружным эмалевым эпителием.[5] Центр петли заполнен клетками звездчатого ретикулума.
Цервикальные петли сохраняются в постоянно растущих резцах грызунов и образуют ниши взрослых стволовых клеток, которые обеспечивают рост зубов.[6]
5)Стадия колокола
На стадии колокола сигнальные молекулы первичного эмалевого узла стимулируют формирование вторичных узлов, которые в свою очередь определяют расположение бугров на поверхности моляров. Происходит это благодаря тому, что клетки эмалевого узла выделяют FGF, который стимулирует окружающий эпителий к пролиферации и одновременно экспрессируют ингибитор клеточного цикла и утрачивают рецепторы к FGF, и потому сами клетки эмалевого узла не делятся.
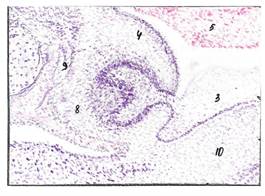
Полость рта эмбриона человека: Окраска гематоксилином и эозином. Ув. х200
1) формирующаяся нижняя челюсть, 2) цилиндрический эпителий; 3) многослойный плоский неороговевающий эпителий; 4) закладка зуба, 5) проксимальный отдел пищеварительной трубки; 6) эмалевый орган; 7) зубной сосочек; 8) зубной мешочек; 9) формирующаяся кость зубной альвеолы; 10) Меккелев хрящ; 11) губа.
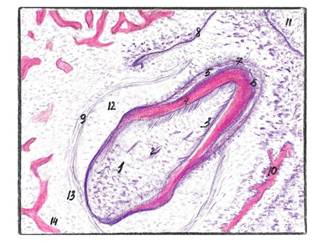
1) пульпа зуба, 2) кровеносные сосуды, 3) дентинобласты, 4) предентин, 5) дентин, 6) эмаль, 7) энамелобласты, 8) зубная пластинка, 9) наружные клетки эмалевого органа, 10) костные трабекулы, 11)многослойный эпителий преддверия ротовой полости, 12) остатки эмалевой пульпы, 13) зубной мешочек, 14)мезенхима
6)Сигнальные молекулы.
Все аспекты морфогенеза зубов регулируются эпителиально-мезенхимальными взаимодействиями, которые опосредуются консервативными сигнальными путями, включая Hedgehog (Hh), Wnt, фактор роста фибробластов (FGF), трансформирующий фактор роста β (TGFb), костный морфогенный белок (BMP) и эктодисплазин (EDA), которые координируют дифференцировку, пролиферацию и апоптоз клеток.
Первым морфогенетическим признаком одонтогенеза является утолщение эпителия, который образует первичный эпителиальный тяж. Его положение регулирует фактор трансмисии OSR-2. Формирование и дальнейшее погружение зубной пластинки в мезенхиму регулируется фактором роста фибробластов FGF.[7]
Сигнальные молекулы, такие как TGFb и BMP, стимулируют клетки мезенхимы к дифференцировке в одонтобласты. WNT10b важна для дифференцировки одонтобластов и регуляцию экспрессии белков дентина, также WNT регулирует обновление зубов у позвоночных. Базальная мембрана, является резервуаром сигнальных молекул, в связи с этим она вовлечена в дифференцировку одонтобластов.
BMP2,4 и TGFb способствуют к дифференцировеке эпителия в амелобласты. SHH из эпителиальных клеток способствует дифференцировке и созреванию амелобластов. WNT3, Эктодисплазин (EDA) и Follistatin являются важнейшими факторами к дифференцировке амелобластов, и эти сигнальные молекулы отсутствуют у мышей. В амелогенезе у мышей участвуют дифференциированные экспрессионные факторы транскрипции Sp6 и Msx2
Заключение
Главной задачей учёных является сохранить человеческие зубы, так как с каждым годом увеличивается число людей с заболеванием зубов, и всё больше людей начинают ставить искусственные импланты. Но импланты не долговечны, как биологические зубы.
В последнее время увеличилось количество исследований по выращиванию зубов из стволовых клеток. Для выращивания полноценного зуба используют 2 компонента: стволовые клетки пульпы и стволовые клетки полости рта. Грандиозного прорыва в данной сфере добились японские учёные по выращивании зубов мышей. Они смогли вырастить резец, но не могли контролировать направленность и скорость развития зуба.
Появляется вопрос: «можно ли использовать эти стволовые клетки на людях?»
После изучения большого количества источников мы пришли к выводу, что стволовые клетки мыши нельзя использовать на человеке. Так как анатомически и гистологически зубы отличаются по своему составу, у мышей зубы растут непрерывно, а также имеются отличия в сигнальных молекулах регулирующих дифференцировку зубов.
На сегодняшний день в данной сфере проводятся обширные исследования по использованию собственных стволовых клеток человека. Для этого используют стволовые клетки пульпы из молочных или недавно удалённых зубов. Для сохранения качества клеточного состава использую разные методы криоконсервации, что может надолго сохранить клетки и при необходимости их разморозить. Остаются лишь вопрос материального обеспечения и сложности проведения данного метода на практике.