

Введение. Для любой водной экосистемы характерны различные процессы массообмена, непрерывно происходящие между её компонентами. Скорости этих процессов определяются множеством условий, из которых важную роль играют абиотические факторы окружающей среды. В настоящее время опубликовано большое количество работ по факториальной экологии, учитывающих влияние отдельных факторов на процессы новообразования и разложения органического вещества. Например, в работах [1, 2] рассматривается зависимость продуктивности водной экосистемы от освещенности и концентрации в воде биогенных элементов. В зарубежных публикациях [3, 4] также описывается влияние температуры, освещенности и концентрации биогенных элементов на величину первичной продукции. Существенным недостатком большинства подобных исследований является то, что в них рассматривается действие одного или нескольких конкретных факторов на процессы массообмена, в то время как в естественной среде наблюдается одновременное действие многих факторов. При этом большие надежды возлагаются на методы биотестирования и на возможности планировать устойчивое (сбалансированное) развитие и коэволюцию природных систем и общества на основе создания моделей функционирования эко- и геосистем. В таких моделях успешность прогнозирования системной динамики во многом зависит от авторского представления о совокупном влиянии естественных и антропогенных факторов на скорости процессов в системе и, прежде всего, продукции и деструкции органического вещества в ней.
По соотношению продукции и деструкции можно понять, какой тип трофии характерен для водоема, какие процессы преобладают в водной экосистеме, способна ли она самоочищаться и справляться с дополнительными нагрузками. В гидроэкологическом мониторинге в настоящее время для ответов на эти вопросы традиционно используется метод светлых и темных склянок для определения величин продукции и деструкции. В конечном счёте соотношение скоростей продукции и деструкции является индикатором продукционно-деструкционного баланса экосистемы водоема, оценка которого представляется еще одной актуальной задачей современности. В большинстве работ для подобных оценок чаще используется покомпонентный анализ, например, в [5] выводы о статусе водоема делаются только на основании величины биогенной нагрузки, содержания биогенных элементов и концентрации хлорофилла. С увеличением числа используемых параметров растет и неоднозначность интерпретации полученных результатов, так как использование разных параметров или большого числа оценочных шкал разных авторов позволяет отнести изучаемый водоём к разным классам. Решением этой проблемы является разработка шкал интегральных показателей, позволяющих выполнить оценку по большому числу параметров или по нескольким группам параметров и нескольким уровням оценивания. Несколько этапов свертки показателей, агрегированных в отдельные группы, содержащие небольшое количество слабо коррелируемых критериев, позволяют устранить мультиколлинеарность и сгладить влияние отдельных сильно отличающихся от основной массы факторов.
Целью исследования является оценка влияния различных факторов на скорости новообразования и разложения органического вещества, а также разработка модели-классификации для интегральной оценки трофности, учитывающей многокритериальность и многоуровневость получения интегральной оценки. Задачи: 1 – учет влияния температуры, освещенности, концентрации биогенных элементов на процессы первичного продуцирования органического вещества в озере Суури и выявление фактора, лимитирующего образование первичной продукции фитопланктоном; 2 – оценка совместного влияния факторов по моделям Либиха, Митчерлиха и ЛМ-модели и сравнение полученных результатов; 3 – анализ натурных наблюдений и моделирование продукционно-деструкционных отношений в озере; 4 – покомпонентная оценка, разработка моделей интегральной оценки трофического статуса водоёма.
Материалы и методы. Количественная оценка влияния температуры, освещенности, биогенных элементов на величину первичной продукции осуществлялось на основе данных, собранных в ходе полевой практики студентов Института наук о Земле СПбГУ (кафедра Гидрологии суши) на учебно-научной базе СПбГУ «Приладожская» в районе п. Кузнечное (Ленинградская область) в июне 2023 года.
Оценка влияния абиотических факторов на скорости трансформации органического вещества в водной экосистеме выполнялась для температуры, освещенности и концентрации биогенных элементов, в число которых вошли минеральный (аммонийный) азот и минеральный фосфор. Связь между температурой воды и максимальной удельной скоростью роста фитопланктона при условии непрерывного освещения устанавливалась для смеси фитопланктона по формуле Eppley [6] или только для диатомового фитопланктона, преобладавшего в озере Суури по формуле Lehman (1975) при условии, что t > tFiопт (tFiопт = 15℃):
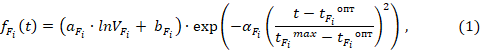
для диатомовых: aFi = – 0,298, αFi = 2,3, VFi = 1500 мкм3, bFi = 4,324.
Оценка влияния освещенности (функция f(I)), осуществлялась на основе формулы Дж. Стила [6]. Также была рассчитана обратная ей функция 1/f(I), которая показывает, во сколько раз недостаток света в воде снижает максимальную удельную скорость роста фитопланктона.
Величина лимитации первичной продукции аммонийным азотом f(N) и минеральным фосфором f(P) выполнялась по модели Михаэлиса-Ментен-Моно [6]. Все расчёты выполнялись на основании средних, максимальных и минимальных значений концентраций, полученных во время проведения суточной станции и в ходе съемки озера. Совместный учет влияния температуры, освещенности, биогенных элементов осуществлялся на основании моделей Либиха (Л-модель), Митчерлиха (М-модель), а также ЛМ-модели (табл. 1).
Таблица 1. Используемые модели для совместного учёта влияния факторов
|
Вид модели |
Вид функции |
|
Л-модель |
|
|
М-модель |
|
|
ЛМ-модель |
|


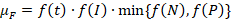
Для изучения скоростей продукции и деструкции использовался метод светлых и темных склянок, которые экспонировались в течение одних суток в свободном от макрофитов открытом районе в центре озера с наибольшими глубинами (около 5 м). Наблюдения проводились в течение 7 суток, при этом использовались ёмкости объема 0,25, 0,5 и 2 л. Сравнение полученных результатов должно было доказать, что величина суточной продукции и деструкции зависит об экспонируемого объема воды.
Продукционно-деструкционное отношение рассчитывалось по модели:
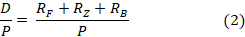
где 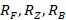 – скорость трат на обмен фитопланктона, зоопланктона и бактерий соответственно (мгО2/л·сут); Р – валовая первичная продукция (мгО2/л·сут). Алгоритм расчета этих скоростей подробно рассмотрен в других наших публикациях, например, в [6].
– скорость трат на обмен фитопланктона, зоопланктона и бактерий соответственно (мгО2/л·сут); Р – валовая первичная продукция (мгО2/л·сут). Алгоритм расчета этих скоростей подробно рассмотрен в других наших публикациях, например, в [6].
Для определения продуцирующей функции озера Суури в июне 2023 года использовался покомпонентный анализ и интегральная оценка. Для этого было отобрано 11 критериев, которые были разделены на три группы, в состав которых вошли гидрофизические, гидрохимические и гидробиологические параметры. Одновременно было введено 5 классов трофности. Для каждого критерия была сформирована оценочная шкала. При выборе шкал мы ориентировались на авторские шкалы или вносили в них небольшие изменения (осреднение, интерполяция, экстраполяция) для их соответствия пяти классам трофности. При разработке шкал для субиндексов и шкалы последнего уровня свертка показателей осуществлялись для левой и правой границ класса внутри каждой группы (первый уровень свертки), а затем полученные значения шкал для субиндексов осреднялись для получения итоговой шкалы интегрального показателя (второй уровень свертки).
Результаты исследования и их обсуждение. Результаты расчётов максимальной удельной скорости роста планктонных водорослей по данным наблюдений за температурой воды в поверхностном горизонте по разным формулам показывает, что и средняя, максимальная и минимальная температуры воды влияют на 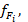 рассчитанную по формуле 2, сильнее, чем по формуле 1 (табл. 2).
рассчитанную по формуле 2, сильнее, чем по формуле 1 (табл. 2).
Таблица 2. Максимальная удельная скорость роста фитопланктона по данным наблюдений на суточной станции в 2023 г.
|
Таксоны фитопланктона |
|
||
|
|
|
|
|
|
1. Диатомовые (по формуле Lehman (1)) |
0,99 |
0,76 |
0,57 |
|
2. Агрегированный (по формуле Eppley [6]) |
2,43 |
2,57 |
2,72 |
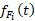
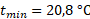
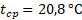
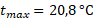
При оценке зависимости удельной скорости роста диатомового фитопланктона от освещенности были получены схожие результаты для суточной станции и съемки озера по средним, максимальным и минимальным величинам прозрачности (табл. 3).
Таблица 3. Зависимость удельной скорости роста диатомовых планктонных водорослей от величины освещенности в 2023 г.
|
Суточная станция |
Съемка озера |
||||
|
Прозрачность, м |
|
|
Прозрачность, м |
|
|
|
|
0,62 |
1,60 |
|
0,63 |
1,59 |
|
|
0,63 |
1,59 |
|
0,62 |
1,59 |
|
|
0,62 |
1,61 |
|
0,63 |
1,59 |
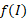

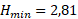
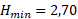


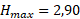
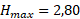
Полученные значения 1/f(I) позволяют сказать, что недостаток освещенности в среднем в 1,60 раза уменьшает максимальную удельную скорость роста диатомового фитопланктона. Такое уменьшение объясняется эффектом самозатенения фитопланктона в верхнем слое толщи воды.
Для расчета зависимости скорости новообразования органического вещества от количества биогенных элементов по модели Михаэлиса-Ментен-Моно была взята средняя для горизонтов 0, 1 и 2 м концентрация аммонийного азота и минерального фосфора. Полученные для суточной станции и съёмки озера результаты отличаются незначительно (табл. 4). Вместе с тем, можно однозначно сказать, что недостаток азота уменьшает величину первичной продукции в среднем в 1,34 раза, а недостаток фосфора – в 2,80 раза, то есть лимитирующим биогенным элементом является минеральный фосфор.
Таблица 4. Зависимость удельной скорости роста диатомовых водорослей от концентрации биогенных элементов
|
Вид работ |
РО4, мгP/л |
NH4, мгN/л |
|
|
|
|
|
Суточная станция |
0,01 |
0,081 |
0,39 |
0,70 |
2,59 |
1,43 |
|
Съёмка озера |
0,01 |
0,133 |
0,33 |
0,79 |
3,00 |
1,26 |
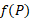
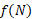


При совместном учёте влияния вышеназванных факторов по трём моделям (табл.1) было получено, что удельная скорость роста агрегированного фитопланктона в большей степени, по сравнению с диатомовыми водорослями, зависит от основных абиотических факторов. В среднем удельная скорость чистого продуцирования фитопланктона  для агрегированного планктона в 3 раза выше, чем для диатомовых водорослей (табл. 5). Также необходимо отметить, что согласно М-модели абиотические факторы в два раза сильнее снижают скорость новообразования органического вещества по сравнению с результатами, полученными по Л-модели.
для агрегированного планктона в 3 раза выше, чем для диатомовых водорослей (табл. 5). Также необходимо отметить, что согласно М-модели абиотические факторы в два раза сильнее снижают скорость новообразования органического вещества по сравнению с результатами, полученными по Л-модели.
Таблица 5. Удельная скорость роста фитопланктона по Л-, М- и ЛМ-моделям
|
Фитопланктон |
|
|
|
|
Агрегированный |
0,92 |
0,43 |
0,58 |
|
Диатомовый |
0,27 |
0,13 |
0,17 |
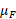 по Л-модели
по Л-моделиДля вычисления D/P-отношения было выполнено 19 серий наблюдений, из них после отбраковки было выбрано 15 серий с экспозицией одни сутки, в том числе 9 серий в склянках объемом 0,25 л и 5 серий в склянках других объемов (0,5 л и 2,0 л). По результатам расчётов в 6 сериях в склянках 0,25 л деструкция превосходила величину продукции; в 3 сериях, наблюдаемых 23-24.06.23, D/P-отношение было меньше единицы, что свидетельствовало об активизации продукционных процессов в озере. Подтвердился вывод о том, что величина первичной продукции зависит от объема склянки. Также исследовалось влияние на результат времени экспозиции проб. Так, при определении первичной продукции в склянках 0,25 л автономным волоконно-оптическим измерителем кислорода «Fibox 4 trace» при экспозиции порядка 1 часа в дневное время (12.00) в разных районах озера величина D/P всегда была меньше 1,0 и в зависимости от района озера различалась в 2-4 раза.
Среднее значение D/P-отношения для объема склянки 0,25 л за все наблюдения в 2023 г. составило 2,79, а для различных объемов проб – 2,55. Из этого следует, что с увеличением объема пробы увеличиваются величины продукции органического вещества. Деструкция также растет, и с повышением температуры воды в водоеме темпы деструкции растут быстрее, чем темпы продукции. В июле 2021 года по 8 сериям наблюдений для склянок объемом 0,25 л D/P-отношение в среднем составило 1,86, причем в 50% случаев оно не превышало 1. За период работ на озере в конце июня – июле 2015 -2023 гг. (в 2020 и 2022 гг. исследования не проводились) было получено, что средняя величина D/P-отношения составила 2,41. Таким образом, данные 2023 г. в целом близки к средним результатам за последние годы.
Для моделирования D/Р-отношения в водоеме по (2) нами использовалась модель, подробно описанная в [6 и 7]. Траты на обмен фитопланктона в этой модели составляют долю от чистой первичной продукции фитопланктона. По (2) было выполнено 30 вариантов расчета D/P-отношения. В вариантах использовались: 1 – разные модели расчета удельных скоростей чистого первичного продуцирования органического вещества фитопланктоном (табл.1); 2 – разные формулы расчета влияния температуры воды на удельную скорость роста фитопланктона, агрегированного – по [6], или с преобладанием в суммарной биомассе одного таксона фитопланктона (диатомовые или зеленые) по формуле (1); 3 – учет возможной недооценки биомассы бактерий в озере, поскольку нами предполагалось, что бактерии, ассоциированные с детритом, составляли примерно 50% от общей биомассы бактерий; 4 – расчеты проводились по данным натурных наблюдений 2023 и 2021 гг.
Результаты сравнения полученных D/Р-отношений по данным натурных наблюдений и по данным моделирования показали, что Л-модель + (диатомовые, формула 1) хорошо описывает полученные в наблюдениях результаты в случае преобладания в составе фитопланктона диатомовых водорослей в 2023 г. (D/Р=2,87) и 2021 г (D/Р=1,95). При этом поправки на недооценку содержания в воде бактерий не требуется. Расчеты по М-модели + (агрегированный фитопланктон по [6]) дают в 2023 г. D/Р=2,02 и 2021 г D/Р=1,76, что вполне согласуется с результатами наблюдений за эти годы, также без поправки на биомассу бактерий или с незначительным её увеличением. Другие варианты дают бóльшие расхождения с полученными по наблюдениям значениями D/Р.
Оценка трофности озера Суури осуществлялась по двум вариантам покомпонентной оценки и двум моделям-классификациям (М1 и М2). В М1 вошло 4 класса трофности и 10 критериев, не разделенных на группы. По результатам покомпонентного анализа №1 (для критериев М1) было получено, что по 3 критериям озеро относится к олиготрофии, по 6 – к мезотрофии, по 1 – к границе между мезо-и эвтрофией. В модель М2 вошло 5 классов трофности, три группы параметров (всего 11 критериев). Процент совпадения критериев моделей составил 80%. В результате покомпонентного анализа №2 (для критериев М2) было получено, что озеро Суури можно отнести к олиготрофному типу водоемов по 3 критериям, к эвтрофному по 1, к мезотрофии 1 по 4, к мезотрофии 2 по 3. Таким образом, в обоих случаях озеро можно признать мезотрофным водоемом в период проведения летних наблюдений.
Интегральная оценка трофности по М1 была выполнена по10 критериям оценивания и 4 классам трофности. Шкала приведена в таблице 6. Было установлено, что для озера Суури в июне 2023 года ИПТ составил 0,230, что позволяет отнести озеро к левой границе мезотрофного класса.
Таблица 6. Шкала интегрального показателя трофности (ИПТ) по М1.
|
Тип трофии |
Олиготрофия |
Мезотрофия |
Эвтрофия |
Гипертрофия |
|
ИПТ |
0,00 – 0,14 |
0,14 – 0,42 |
0,42 – 0,77 |
0,77 – 1,00 |
В модель М2 (рис.1, табл.7) вошло 11 критериев, 5 классов трофности (класс мезотрофных вод был разбит на два: мезотрофия 1 и мезотрофия 2), 2 уровня свертки и 3 группы параметров, учитывающих гидрохимические, гидрофизические и гидробиологические параметры. Свертки показателей на всех уровнях осуществлялись для равновесомых условий учета критериев.
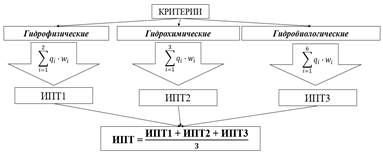
Рис. 1. Схема расчёта интегрального показателя трофности по М2.
Таблица 7. Классификация для интегральной оценки трофности водоема по М2.
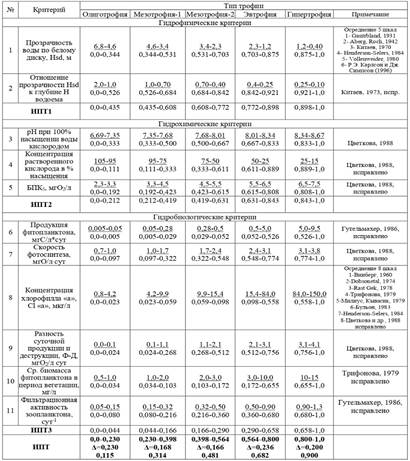
Примечание: в числителе – абсолютные значения параметров, в знаменателе – нормированные значения. В шкале ИПТ 3 строки: в первой строке – левое и правое значение ИПТ для каждого класса; Δ – ширина класса, в последней строке – среднее значение ИП для класса.
В ходе вычисления интегрального показателя трофности было установлено, что по гидрофизическим параметрам озеро относится к мезотрофии-2 (ИПТ1 составляет 0,625), по гидрохимическим и гидробиологическим – к мезотрофии-1 (ИПТ2 и ИПТ3 составляют 0,311 и 0,088 соответственно). Интегральный показатель трофности составил 0,341 (мезотрофия-1, ближе к середине класса). Сравнивая результаты, полученные по М1 и М2, можно отметить, что по обеим шкалам рассчитанный ИПТ для озера Суури соответствует левой части мезотрофного класса трофности (рис. 2).

Рисунок 2. Сравнение результатов интегральной оценки трофности оз.Суури по вариантам М1 и М2
Заключение. При оценке влияния факторов на продуктивность водной экосистемы было установлено, что недостаток освещенности снижает скорость образования первичной продукции в 1,6 раза, а лимитирующим биогеном является минеральный фосфор. При совместном учете факторов по трём моделям было получено, что для диатомового фитопланктона удельная скорость роста меньше, чем для агрегированного. Кроме того, по М-модели скорость новообразования органического вещества получается наименьшая, а по Л-модели наибольшая.
По наблюдениям получено, что в среднем в июне-июле скорость деструкции органического вещества превышает скорость продукции в озере в 2,55 раза. С увеличением объема пробы увеличиваются величины продукции органического вещества. Деструкция также растет, и с повышением температуры воды в водоеме темпы деструкции начинают расти быстрее, чем темпы продукции. Результаты моделирования реально описывают наблюденные D/Р-отношения при использовании Л-модели для диатомового фитопланктона и М-модели для агрегированных микроводорослей. В обоих случаях поправки на возможный недоучёт трат на обмен бактерий не требуются.
Покомпонентная оценка трофности позволила сделать вывод о том, что по большинству параметров озеро Суури относится к мезотрофному типу. В ходе интегральной оценки по М1 с использованием 10 критериев был получен ИПТ = 0,23, что позволило отнести озеро к левой части мезотрофного класса. Аналогичные результаты наблюдались при использовании М2 по 11 критериям, ИПТ = 0,34, что соответствует левой части мезотрофного класса. Близость полученных по М1 и М2 результатов убеждает в том, что незначительное расхождение в количестве и составе выбранных оценочных критериев (в пределах 20%) не повлияло на общую оценку трофности водоема.
Работа выполнена при финансовой поддержке Российского научного фонда, грант № 23-27-10011, и при финансовой поддержке Санкт-Петербургского научного фонда.