Введение
Антибиотики произвели революцию в современной медицине, позволив добиться значительного прогресса в качестве и продолжительности жизни. Всемирная организация здравоохранения в 2014 году предупредила, что мы вступаем в «эру после антибиотиков». Новый кризис, с которым сегодня сталкивается мир — это устойчивость к антибиотикам. Генетические модификации в бактериях привели к состоянию, при котором патогенные микроорганизмы стали устойчивыми к доступным антимикробным агентам. Риск резистентности также был ускорен из-за нерационального приема антибиотиков. Наиболее распространенным примером тому являются люди, страдающие простудой и кашлем.
В последнее время не было разработано ни одного нового класса антибиотиков, а с 1962 года было создано только два класса. Антибиотики были открыты ранее путем скрининга почвенных микроорганизмов, которые являются основным источником потенциальных новых веществ с антимикробной активностью, однако 99% бактерий не могут быть выращены при современных методах. Исследователи из лаборатории Льюиса и Раба Эпштейна разработали новую технику под названием iChip, которая позволяет исследовать некультивируемые бактерии, поскольку они растут в естественной для них среде и выделять производимый ими антибиотик, а затем идентифицировать его.
В пресс-релизе от 7 января 2015 года NovoBiotic Pharmaceuticals объявила об открытии тейксобактина, нового антибиотика из ранее некультивированной и неописанной почвенной бактерии, принадлежащей к β-протеобактериям, названной Eleftheria terra. Тейксобактин оказался первым представителем нового класса антибиотиков со значительной активностью в отношении патогенных штаммов, таких как ванкомицин устойчивых энтерококков (VRE) и метициллин устойчивых Staphylococcus aureus (MRSA). Тейксобактин связывается с предшественником пептидогликана и тейхоевых кислот, являющиеся компонентами клеточной стенки и, следовательно, ингибирует биосинтез клеточной стенки бактерий.
Цели работы: обосновать рациональность будущего использования тейксобактина.
Задачи:
1. Описать возможности технологии iChip.
2. Обосновать высокую антибактериальную активность тейксобактина исходя из структуры молекулы и механизма действия.
3. Обозначить спектр активности и потенциал развития резистентности.
1. Историческая справка
В 1943 году Сельман Ваксман и Альберт Штатц сооткрыли аминогликозидный антибиотик стрептомицин. Это был первый эффективный антибиотик для лечения туберкулеза, используемый по сей день. Вслед за этим знаменательным событием Ваксман получил в 1952 году Нобелевскую премию за «гениальные, систематические и успешные исследования почвенных микробов, которые привели к открытию стрептомицина». В отличии от ранее случайного открытия пенициллина Александром Флемингом, который заметил, что загрязнение плесенью в культуре чашки Петри препятствовало росту патогенных бактерий. Ваксман и его команда разработали платформу для выявления новых антибиотиков: она позволила проводить скрининг полученных из почвы актиномицетов на антибактериальную активность против микроорганизмов в чашках с агаром. Используя этот метод в 1940-х годах, Ваксман и его ученики выделили более пятнадцати антибиотиков, в том числе стрептомицин и неомицин [6].
В течение следующих десятилетий на основании метода скрининга почвенных микроорганизмов были открыты основные классы антибиотиков, например макролиды, тетрациклины и рифамицины. К сожалению, этот метод прекратил давать результаты в середине шестидесятых годов, поскольку область исследования была ограничена лишь 1% разнообразия микробов во внешней среде, так как остальные 99% до недавних лет были некультивируемыми и недоступными для изучения, так как они не способны расти в лабораторных условиях [10].
В настоящее время микробиологические культуры выращиваются преимущественно в твердом или жидком агаре. В течение многих лет техника выращивания бактерий в лаборатории оставалась неизменной. Подавляющее большинство микробов, видимых в микроскопе, в настоящее время не может быть выращено в лабораториях, поскольку некоторые бактерии могут фактически быть нежизнеспособными в этих условиях, так как микроорганизмы приспособились к процветанию в условиях их собственной родной среды.
В целях расширения доступа к большему разнообразию микроорганизмов был разработан новый метод культивирования в собственной среде обитания под названием iChip (изолирующий чип), который уже выделил множество новых организмов, а также тейксобактин — антибиотик со значительной активностью против грамположительных бактерий, в том числе, обладающих резистентностью к современным антибиотикам. Однако этот антибиотик все еще нуждается в клинических испытаниях. Сообщалось, что тейксобактин не эффективен в отношении грамотрицательных бактерий, что обусловлено строением их клеточной стенки и механизмом действия тейксобактина.
2. Принцип работы устройства iChip
Исследователи из Северо-Восточного университета и Novobiotic Pharmaceuticals разработали революционную технологию, в которой используется многоканальное устройство, называемое iChip, для выделения и выращивания некультурных бактерий в их естественной среде. Эта техника позволила выращивать большее количество микроорганизмов по сравнению с традиционной чашкой Петри (50% против 1%) [8].
IChip представляет собой сборку пластиковых пластин и мембран для захвата микроорганизмов окружающей среды (Приложение 1) [14]. По сути, он состоит из пластиковых пластин, которые содержат около сотни отверстий, каждое из которых образует очень маленькую диффузионную камеру, в которую помещается только один микроорганизм. Чип содержит 96 миниатюрных диффузионных камер, которые обеспечивают прохождение питательных веществ и метаболитов через полупроницаемую мембрану, удерживая при этом внутри камер бактериальные клетки.
Для выделения микроорганизмов образцы собираются из, например, почвы или морской воды, и смешиваются с жидким агаром. Предварительно образец почвы разбавляется до такой степени, что бы в каждую из камер iChip попадала примерно одна бактериальная клетка. Когда центральная пластина погружена в суспензию клеток в расплавленном агаре, сквозные отверстия заполняются небольшим объемом этой суспензии, которая затвердевает в виде небольших агаровых пробок.
Затем устройство покрывают двумя полупроницаемыми мембранами (размер пор 0,03 мкм) для обеспечения диффузии питательных веществ и факторов роста, и помещают обратно в почву, из которой была взята проба. Через некоторое время, выросшую колонию можно перенести в лабораторию и исследовать.
Этот метод скрининга значительно облегчит обнаружение новых антибиотиков, поскольку он позволяет соединения из микроорганизмов окружающей среды, которые не растут в нормальных лабораторных условиях [1]. Устройство позволило вырастить значительное количество бактериальных колоний по сравнению с чашкой с агаром.
Экстракты из 10000 изолятов, полученных в результате роста в iChip бактерий, были подвергнуты скринингу на антимикробную активность на пластинках покрытых S. aureus. Это привело к обнаружению нового вида грамотрицательного микроорганизма, условно названного Eleftheria terrae из вида b-протеобактерий, который продуцировал соединение, проявившее высокую антибактериальную активность [14].
3. Структура молекулы тейксобактина
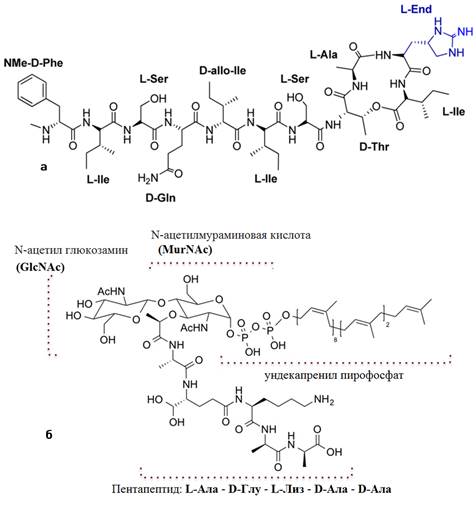
Структура была определена посредством обширного анализа методами жидкостной хроматографии, масс-спектрометрии и методами ядерного магнитного резонанса (ЯМР). Эти анализы выявили необычный депсипептид, содержащий 11 аминокислот, связаные примерно 10 пептидными связями (Приложение 2-а) [6].
Молекулу можно условно разделить на головную и боковую цепи. Боковая цепь состоит из 7 аминокислот: N-метил-D-фенилаланин, D-глутамин, два L-изолейцина и один D-изолейцин, два L-серина [16]. Головная цепь представляет собой макролактоновое кольцо, состоящее из четырех аминокислот: D-треонина, L-аланина, L-изолейцина и L-алло-эндурацидина — непротеиногенной аминокислоты, которая была впервые идентифицирована как аминокислотный компонент сильнодействующих депсипептидных антибиотиков, выделенных из Streptomyces hygroscopicus LL-AC98 в 2002 году [19].
4. Биосинтез пептидогликана
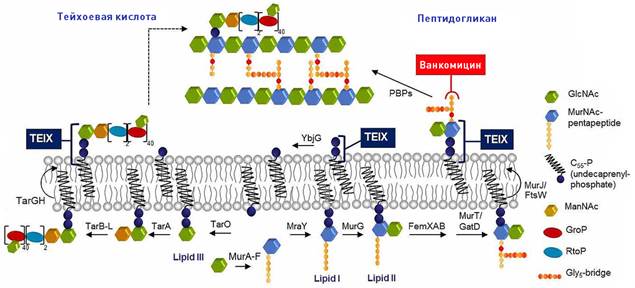
Бактериальная клеточная стенка жизненно необходима для бактерий, поскольку она противодействует осмотическому давлению, которое возникает между цитозолем клетки и ее окружающей средой. Стенка состоит из слоев пептидогликана, сшитых линейными гликановыми цепями [18]. Пептидогликан, основной компонент клеточной стенки грамположительных бактерий, синтезируется из липида II — строительного блока, уникального для бактериальных клеток. Ферменты, сшивающие пептидогликана являются мишенью для β-лактамных и гликопептидных антибиотиков.
Липид II — предшественник пептидогликана, кроме липидного компонента и пирофосфата, содержит основные строительные блоки пептидогликана: N-ацетилмураминовую кислоту, N-ацетилглюкозамин и пентапептид (L-Ala-D-Glu-L-Lys / DAP-D-Ala-D-Ala) (Приложение 3) [13].
У грамположительных бактерий тейхоевая кислота (ТА) также является основным компонентом клеточной стенки. TA включает стеночную тейхоевую кислоту (WTA), связанную с пептидогликаном, или липотейхоевую кислоту (LTA), закрепленную в цитоплазматической мембране. TA играет важную роль в физиологии бактерий [18], а ферменты биосинтеза тейхоевой кислоты являются важной мишенью для антибиотиков. Недавно было показано, что ингибирование биосинтеза тейхоевой кислоты может восстановить чувствительность к метициллину у метициллин-резистентного золотистого стафилококка (MRSA) [9].
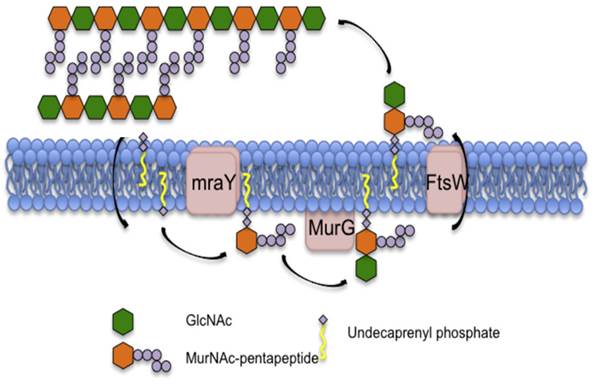
Синтез липида II начинается на цитозольной стороне мембраны, где встроенный в мембрану фермент MraY связывает пентапептид UDP-N-ацетил-мураминовой кислоты (UDP-MurNAc-pp) с липидом-ундекапренилфосфатом, образующим липид I. Затем трансфераза MurG связывает UDP-N-ацетилглюкозамин (UDP-GlcNAc) с мурамоильной частью липида I с образованием липида II. Затем FtsW «переворачивает» липид II на другую сторону бактериальной мембраны, после чего белки, связывающие пенициллин (PBP), включают липид II в растущую сеть пептидогликана. Остаточный ундекапренилпирофосфат затем дефосфорилируется до монофосфата и переворачивается обратно в цитозольный сайт для повторного использования в цикле синтеза липида II(Приложение 2-б) [7].
MurNAc-пентапептид связан с ундекапренилфосфатом с помощью mraY. Затем GlcNAc связывается с липидом I с помощью MurG с образованием липида II. Затем предшественник пептидогликана переворачивается в периплазматическое пространство с помощью ftsW и включается в пептидогликан пептидами, связывающими пенициллин (PBP).
Жизнеспособность липида II как мишени для гликопептидных антибиотиков была ясно продемонстрирована ванкомицином, который впервые был выделен из почвенных бактерий Streptomyces orientalis в 1953 году Эли Лили. Начиная с его введения в клинику, ванкомицин остается единственным антибиотиком, мишенью которого является липид II, и, к тому же, является последним средством лечения против многих устойчивых к пенициллину и метициллину грамположительных бактерий. На ранних испытаниях ванкомицина исследователи не смогли выделить бактерии, устойчивые к ванкомицину [13]. Однако после более чем 30 лет клинического применения резистентность к ванкомицину все же возникла.
В настоящее время известно шесть различных генотипов резистентных к ванкомицину бактерий, обозначаемых как кластеры генов VanA-VanG. Бактерии с резистентностью к VanA-, VanB- и VanD-типам обладают измененной формой липида, где связывающий ванкомицин мотив D-Ala-D-Ala заменен на D-Ala-D-Lac [4]. Другие устойчивые к ванкомицину микроорганизмы вместо этого используют D-Ala-D-Ser, хотя это приводит к менее серьезной устойчивости. Связывание ванкомицина с D-Ala-D-Ala фрагментом липида II хорошо изучено и обусловлено образованием пяти водородных связей. Замена терминального D-аланина на D-Lac или D-Ser позволяет ванкомицину образовывать только четыре водородные связи, что приводит к почти 1000-кратной потере сродства к его цели [12]. Этот способ обхода связывания липида II, вероятно, основан на механизме самоиммунитета штамма-продуцента или бактерий, которые естественным образом несут этот фрагмент, таких как Lactobacillus Casei и Lactobacillus Plantarum.
5. Особенности связывания тейксобактина с предшественником пептидогликана
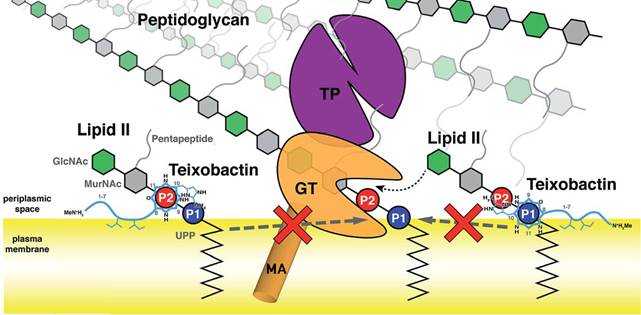
Были обнаружены две основные конформации связи липид II-тейксобактин, каждая из которых демонстрирует характерное связывание пирофосфата липида II амидами головной цепи тейксобактина (Txb) вблизи С-концевого циклодепсипептидного (D-Thr8-Ile11) кольца (Приложение 4) [11]. Кроме того, в связывании также участвует гидроксильная группа боковой цепи Ser7 и вторичное фосфатное связывание, обеспечиваемое головной цепью (L-алло-эндурацидин). Интересно, что эти конформации различаются заменой двух групп доноров водородных связей, которые координируют две фосфатные группы липида II, что приводит к противоположным ориентациям связывания липида II [5].
Кроме того, остатки D-allo-Ile5 и Ile6 служат в качестве мембранных якорей в обеих конформациях Txb, независимо от детальных фосфат-связывающих взаимодействий вблизи депсипептидного кольца. Роль гидрофобных остатков в активности Txb, в первую очередь, заключается в его мембранном закреплении и, кроме того, в обеспечении неполярных взаимодействий с хвостом липида II.
В обеих конформациях связывание липида II достигается в основном за счет координации фосфатов вблизи С-концевого циклодепсипептидного кольца, в то время как два гидрофобных остатка в середине пептидной цепи служат в качестве якоря мембраны. Не наблюдается значительного взаимодействия между Txb и дисахаридом или пентапептидом липида II. Хотя гидрофобные остатки в Txb необходимы для его активности, это требование важно для закрепления Txb на поверхности мембраны, а не для стимулирования специфического связывания с липидом II, включающего гидрофобные взаимодействия. Таким образом, существенными для активности Txb представляются как специфическое связывание с фосфатом липида II, так и неспецифическое связывание с поверхностью мембраны.
6. Механизм действия
Способность тейксобактина одновременно ингибировать биосинтез пептидогликана и тейхоевой кислоты вызывает синергические эффекты, приводящие к повреждению клеточной стенки, делокализации аутолизинов и последующему лизису и гибели клеток. Кроме того, тейксобактин не связывает зрелый пептидогликан и, следовательно, способен эффективно воздействовать на штаммы ванкомицина-промежуточного S. aureus (VISA), которые имеют повышенную плотность клеточной стенки.
Было показано, что механизм действия тейксобактина основан на связывании двух важных строительных блоков бактериальной клеточной стенки: липид II (основной предшественник пептидогликана), и липид III (предшественник тейхоевых кислот) [7]. Тейхоевые кислоты играют важную роль в предотвращении неконтролируемого гидролиза пептидогликана, поэтому ингибирование синтеза тейхоевых кислот может способствовать делокализации аутолизинов. Липид II также является мишенью для гликопептидного антибиотика ванкомицина, но тейксобактин связывается с другой его областью и поэтому активен против устойчивых к ванкомицину энтерококков (VRE), которые имеют модифицированный липид II. Таким образом, он одновременно ингибирует биосинтез пептидогликана и тейхоевой кислоты, вызывая синергетический эффект, приводящий к повреждению клеточной стенки.
Липид II, предшественник пептидогликана, синтезируется в цитоплазме и переворачивается на поверхность внутренней мембраны с помощью MurJ48 или FtsW49 (Приложени 5) [7]. Липид III, предшественник тейхоевой кислоты в стенке (WTA), аналогичным образом образуется внутри клетки, и липид-связанные предшественники WTA транслоцируются через цитоплазматическую мембрану с помощью ABC-транспортера TarGH50 [7].
Биосинтез липида II варьируется внутри пентапептида в третьем положении между грамположительными бактериями (лизин) и грамотрицательными бактериями (диаминопимелиновая кислота (DAP)) [2]. Mикoбактерии spp. обладают DAP и могут дополнительно модифицировать N-ацетил- в N-гликолилмурамовую кислоту. Было обнаружено и несколько других модификаций пептидного стебля или гликановых цепей. Предполагается, что тейксобактин взаимодействует с пирофосфатом липида II и, таким образом, может связывать как грамположительные, так и грамотрицательные варианты липида II, независимо от модификаций сахаров или пептидов.
Ванкомицин также нацелен на липид II. Однако, принимая во внимание то, что тейксобактин проявил активность против ванкомицин-резистентного энтерококка (VRE), его связывание происходит через другую область по сравнению с таковой у ванкомицина. В этом отношении биохимические анализы показали, что тейксобактин связывает пирофосфат и первый сахарный фрагмент, присутствующий как в липиде II, так и в липиде III [7]. Кроме того, тейксобактин, в отличие от ванкомицина, не связывает зрелый пептидогликан, что обеспечивает эффективное уничтожение бактерий с повышенной плотностью клеточной стенки, таких как ванкомицин-промежуточный Staphylococcus aureus (VISA), против которого ванкомицин неэффективен.
Ингибирование биосинтеза тейхоевой кислоты клеточной стенки приводит к делокализации аутолизинов и лизису клеток. Было высказано предположение, что тейхоевые кислоты контролируют связывание аутолизина с пептидогликаном по принципу исключения, при котором взаимодействие тейхоевой кислоты клеточной стенки с пептидогликаном ингибирует связывание аутолизина [15]. Следовательно, отсутствие тейхоевой кислоты в перегородке способствует соответствующей делокализации аутолизинов [3].
Тейксобактин-индуцированный лизис зависит от аутолизина Atl. Известно, что индуцированный β-лактамом лизис опосредуется Atl, основным аутолизином клеточной стенки S. aureus. Обработка тейксобактином приводила к делокализации Atl вследствие ингибирования биосинтеза WTA клетки, что объясняет интенсивную литическую способность тейксобактина [15].
7. Спектр активности
Тейксобактин обладает активностью против грамположительных микробов, включая ванкомицин-устойчивые энтерококки (VRE) и метициллин-устойчивый Staphylococcus aureus (MRSA) [6]. Тейксобактин также был эффективен в качестве однократной дозы для значительного снижения количества бактерий на мышиной модели сепсиса MRSA и пневмококковой пневмонии. На мышах он продемонстрировал активность против нескольких устойчивых штаммов бактерий, таких как Streptococcus pneumonia и Mycobacterium tuberculosis. Он также эффективен против Clostridium difficile, M. tuberculosis и потенциально может использоваться для лечения туберкулеза.
Линг и соавт. в своих исследованиях in vitro показали, что тейксобактин был высокоэффективен против Bacillus anthracis и C. difficile при минимальной ингибирующей концентрации (MIC) 5 и 20 мг / мл [7]. Следует заметить, что в случае M. tuberculosis или S. aureus резистентные штаммы не были обнаружены при посеве с низкой дозой тейксобактина.
При исследовании на лабораторных животных, соединение сохранило свою активность в присутствии сыворотки крови, имело хорошую микросомальную стабильность и низкую токсичность. Фармакокинетические параметры, определенные после внутривенных инъекций одной дозы 20 мг на кг у мышей были благоприятными, поскольку уровень соединения в сыворотке поддерживали выше значения MIC в течение 4 ч. Затем проводили исследование эффективности на мышах: они были внутрибрюшинно инфицированы устойчивым к метициллину S. aureus (MRSA) в дозе, приводящей к 90% смерти. Через час после заражения вводили тейксобактин внутривенно в однократных дозах от 1 до 20 мг на кг. Все животные выжили, и в последующем эксперименте было определено, что PD50 (защитная доза, при которой выживает половина животных) составляет 0,2 мг/кг, что выгодно отличается от 2,75 мг/кг PD50 ванкомицина — основного антибиотика, используемого для лечения MRSA [7].
8. Пути развития резистентности
Мутанты S. aureus или M. tuberculosis, устойчивые к тейксобактину, не продуцировались даже при посеве на среду с низкой дозой соединения. Последовательный пассаж S. aureus в присутствии сублетальной дозы тейксобактина в течение 27 дней также не давал устойчивых мутантов. Обычно это означает, что вещество обладает неспецифическим механизмом действия с сопутствующей токсичностью [7]. Однако тейксобактин не обладал токсичностью в отношении клеток млекопитающих, не проявлял гемолитической активности и не связывал ни ДНК, ни белок. Он связывается с консервативной областью ундекапренил-PP-сахара, которая, как известно, не модифицируется. Мишенями большинства других антибиотиков являются белки, и бактерии становятся устойчивыми в результате мутаций генов, кодирующих эти белки. Но нацеливаясь на липиды вместо белков, бактериям может потребоваться больше времени для развития резистентности. Ванкомицин обладает схожим механизмом действия, и для развития резистентности потребовалось 30 лет.
Микроорганизмы, производящие антибиотик, а иногда и его близкие микробные соседи, устойчивы к антибиотику, так как не имеют для него мишеней либо обладают генами, обуславливающими устойчивость к нему. Для продуцирующего микроорганизма это необходимо, иначе собственно произведенный антибиотик будет действовать на него самого. Eleftheria terrae является грамотрицательной бактерией и, следовательно, не имеет мишени для тейксобактина, который вызывает лизис грамположительных бактерий. Живущие по соседству в той же среде микроорганизмы могут быть устойчивыми, поскольку имеют соответствующие гены устойчивости так, что обуславливает их выживаемость в той же нише, что и продуцирующий микроорганизм.
«Естественные» гены устойчивости к антибиотикам могут быть переданы в патогенные бактерии. Например, ген, кодирующий один из наиболее распространенных механизмов устойчивости к антибиотикам — бета-лактамаза расширенного спектра (ESBL), был получен от бактерии из внешней среды [20]. Таков наиболее вероятный путь развития устойчивости, если это произойдет с тейксобактином. Однако это маловероятно, поскольку патогенные бактерии должны были бы смешиваться с устойчивыми к тейксобактину почвенными бактериями. Тем не менее, чтобы быть уверенным в том, что резистентность к тейксобактину маловероятна при применении в медицине, бактерии, выделенные из той же среды, что и Eleftheria terrae, должны быть подвергнуты скринингу на выявление генов, обеспечивающих устойчивость к тейксобактину.
Выводы
1. Новая методика культивирования с использованием изолирующего чипа (iChip) может обеспечить доступ к неиспользованному резервуару антибиотиков, вырабатываемых некультивируемыми бактериями, поскольку они могут расти в естественных для них условиях.
2. Тейксобактин представляет собой депсипептид, состоящий из 11 аминокислот, четверо из которых образуют макролактоновое кольцо. Механизм действия тейксобактина основан на ингибировании синтеза клеточной стенки путем связывания предшественников пептидогликана и тейхоевых кислот (липид II и липид III соответственно) и дальнейшем лизисе клетки.
3. Тейксобактин активен в отношении грамположительных бактерий, включая ванкомицин-устойчивые энтерококки и метициллин-устойчивый золотистый стафилококк. Так как мишенью тейксобактина являются не ферменты, а несколько высококонсервативных фрагментов предшественников компонентов клеточной стенки, для возникновения мутаций, из-за которых может возникнуть устойчивость к тейксобактину понадобится довольно долгое время.
Заключение
Чтобы тейксобактин (как и любое новое соединение с антимикробной активностью) стал лекарством для лечения инфекций у людей, необходимо провести клинические испытания, чтобы убедиться, что препарат безопасен, хорошо переносится и эффективен у пациентов. Для этого необходимо будет провести полные токсикологические тесты, чтобы убедиться, что после введения тейксобактина нет побочных реакций или взаимодействий между лекарственными средствами. Также необходимо подобрать лекарственную форму так, чтобы антибиотик оставался активным in vivo в клинически значимых местах инфекции. Будет ли он полностью разработан в качестве нового препарата, еще неизвестно, не в последнюю очередь потому, что сомнительно, требуется ли больше лекарств против грамположительных бактерий. Однако, поскольку тейксобактин активен в отношении M. tuberculosis, он может предложить вариант нового лечения для пациентов с туберкулезом. Тейксобактин также может ответить требованиям для одобрения FDA в рамках системы квалифицированных продуктов для инфекционных заболеваний (QIDP), как это предусмотрено в Законе США о создании стимулов для антибиотиков (GAIN), поэтому он может быть быстро лицензирован. Даже если сам тейксобактин не появится на полках аптек, он является первым из серии новых антибиотиков в своем классе.
Приложение 1. iChip: a – центральная пластина, погруженная в суспензию клеток в расплавленном агаре; б – отдельные микроорганизмы в каждой из диффузионных камер; в – полупроницаемые мембраны, отделяющие пластину от окружающей среды и расположенные сверху и снизу от нее, и две опорные боковые панели, которые имеют с центральной пластиной соответствующие сквозные отверстия. [14]

Приложение 2. Химическая структура тейксобактина (а) и липида II (б) [6], [7].
Приложение 3. Схематический обзор пути биосинтеза пептидогликана [13].
Приложение 4. Схематическое изображение двух ориентаций комплексов тейксобактин-липид II на поверхности мембраны и механизм ингибирования биосинтеза пептидогликана. Прототип пенициллинсвязывающего белка показан как комбинация мембранного якоря (MA, оранжевый цилиндр), гликозилтрансферазного домена (GT, оранжевый) и транспептидазного домена (TP, фиолетовый) [11].
Приложение 5. Модель механизма действия тейксобактина, ингибирование синтеза клеточной стенки тейксобактином. Тейксобактин (TEIX) образует стехиометрический комплекс с липидом II и липидом III. Связывание этих строительных блоков одновременно прерывает биосинтез пептидогликана (справа) и тейхоевой кислоты (слева). Связывание с несколькими мишенями в клеточных стенках препятствует образованию функциональной клеточной оболочки. [7]