

Введение. Влияние экзогенных и эндогенных факторов на развитие органов и систем получило новый импульс к исследованиям после серии работ, позволившей выдвинуть гипотезу эмбриогенетической этиологии ряда хронических заболеваний, иммунопатологий и эндокринопатий [1, 2, 3]. Исходя из этой гипотезы, и пренатальные и перинатальные события могут выступать как основа для функционального и структурного развития организмов. В связи с этим можно считать , что перинатальные события могут запоминаться организмом, который развивается, посредством эмбрионального и неонатального импринтинга.
На каждый фенотип клетки может влиять её внутренняя и внешняя среда. В организме человека в процессе развития имеется около 200 видов специализированных клеток, на которые влияют эндогенные и экзогенные стероидные гормоны [5].
Стероидные гормоны модулируют широкий спектр клеточных и физиологических реакций.
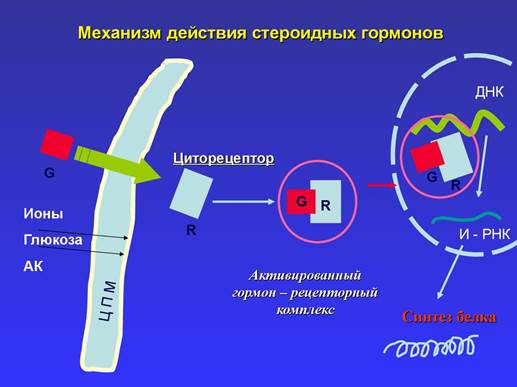
Рис 1: Механизм действия стероидных гормонов
Во время ранних стадий беременности гормоны стероидной природы могут посылать сигналы, которые способствуют успешному внедрению эмбриона в заднюю маточную стенку, и оказывать регулирующее действие на пролиферацию, транскрипцию и дифференцировку генов. Более поздние сроки беременности характеризуются тем, что стероиды влияют на поддержание развития и роста плода посредством модуляции метаболических процессов во время эмбрионального развития и закладки органов. Стероидные гормоны оказывают главенствующее влияние на регулирование сроков родов. Разные гормоны стероидной природы экспрессируются во время различных этапов внутриутробного развития. Временная и пространственная экспрессия стероидов оказывает наиболее важное влияние на рост и развитие плода. Из массы гормонов, которые обнаружены в организме матери во время беременности, эстрогены вызывают максимальный интерес, исходя из того, что они обнаруживаются на всех этапах беременности и оказывают регулирующее и модулирующее влияние на многие процессы, происходящие в матке.
Эстрогены синтезируются в плаценте, яичниках и корковом веществе надпочечников. В период беременности плацента является наиболее важным местом для синтеза эстрогенов, не смотря на это, синтез плацентарного эстрогена возможен лишь при участии коры надпочечников как матери, так и ребенка. Это обусловлено тем, что плацента не способна к самостоятельной выработке андрогенного C19 стероида (дегидроэпиандростерон, ДГЭА и его сульфатированная форма ДГЭАС),который является основой для плацентарного эстрогена [6, 8, 9]. Фетальная железа надпочечников участвует в выработке большого количества побочных веществ, которые используются в синтезе плацентарного эстрогена [10]. Исходя их этого, правильное развитие и функционирование коркового вещества надпочечников является важным для синтеза плацентарных стероидов (например, эстрогена, кортизола и альдостерона) созревания плода и перинатальной выживаемости [11].
Анализ и обсуждение. Надпочечники развиваются из двух различных эмбриональных зачатков. Мозговое вещество происходит из клеток нервного гребня, тогда как кора имеет мезодермальное происхождение [11]. Появление надпочечников в виде адреногонадного зачатка (АГЗ) происходит через 28-30 дней после зачатия у людей (9-й день эмбрионального развития у мышей) и характеризуется экспрессией стероидогенного фактора 1 (SF1), который принадлежит к семейству ядерных рецепторов, факторов внутриклеточной транскрипции, и кодируется NR5A1 геном. Этот фактор необходим для развития надпочечников и стероидогенеза [12]. Как парный орган АГЗ впервые появляется как утолщение целомического эпителия между урогенитальным гребнем и дорзальной брыжейкой. Каждый АГЗ содержит смешанную популяцию клеток: адренокорикальных и соматических гонадных прогениторных клеток. SF1- положительные АГЗ-клетки затем отслаиваются от эпителия и внедряются в мезенхиму промежуточной мезодермы. [13]. После расслаивания большинство клеток АГЗ мигрируют дорзолатерально для образования первичных гонад. Субпопуляция клеток АГЗ, которые экспрессируют более высокие уровни SF1, мигрируют дорзомедиально для формирования первичных надпочечников, так называемая надпочечниковая фетальная зона (fetal zone, FZ). На 48-й день эмбрионального развития у людей (11,5-13,5 день у мышей) клетки нервного гребня мигрируют в развивающуюся FZ. [14]. Эти клетки сохраняются в виде отдельных островков, разбросанных по всему эмбриональному надпочечнику вплоть до рождения, и, в конечном счёте, сливаются и дифференцируются в продуцирующие катехоламин хромаффинные клетки [15]. Далее надпочечник начинает отделяться от окружающей мезенхимы и инкапсулируется с образованием волокнистого слоя над развивающимися кортикальными клетками. Это процесс в основном завершается к 52 дню эмбрионального развития у людей (на 14,5 у мышей) [16].
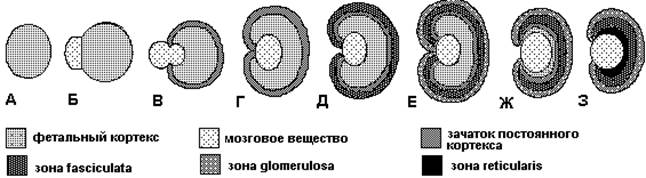
Рис 2: Стадии развития надпочечников
Адренокортикальные и хромаффинные клетки тесно связаны во время эмбрионального и постнатального развития. Глюкокортикоиды надпочечников играют важную роль в продукции гормонов хромаффинными клетками, тем самым регулируя экспрессию N-метилтрансферазы фенилэтаноламина (PNMT),что обусловливает то, что адреналин выступает в роли доминирующего катехоламина, который продуцируется в мозговом веществе надпочечниковой железы в период постнатального развития. [17].
После инкапсулирования кора надпочечников эмбриона быстро расширяется. Надпочечник плода человека является одним из крупнейших органов на этом сроке (0,2% от общей массы тела и почти размером с почку), причем 80% железы состоит из клеток фетальной зоны. Эти крупные стероидогенные клетки (20-50 мм) имеют высокое цитоплазма/ядерное соотношение и устойчиво экспрессируют цитохром P450 17альфа (CYP17), бифункциональный фермент с 17-гидроксилазу и 17,20-лиазу, превращая прегненолон в дегидроэпиандростерон (ДГЭА). Из-за высокой активности CYP17 на этом этапе кора надпочечников плода человека вырабатывает большое количество ДГЭА и ДГЭАС, которые затем преобразуются плацентой в эстрогены для поддержания нормальной беременности. В это же время большое количество других сульфатированных дельта-5-стероидов, включая сульфат прегненолона и сульфат 17α-гидроксипрегненолона, также продуцируются клетками фетальной зоны. К 8- й неделе беременности появляются новые адренокортикальные клетки между капсулой и фетальной зоной, образуя дефинитивную зону, которая позже развивается в кору надпочечников взрослого. Дефинитивная зона состоит из SF1-позитивных, плотно упакованных базофильных клеток, расположенных в узкой полосе клеточных кластеров [18, 19]. Небольшие по размеру (10-20 мкм), клетки дефинитивной зоны активно пролиферируют. По мере развития беременности внутренние клетки FZ образуют арочные тяжи с пальцеобразными колоннами клеток, достигающими внешнего края фетальной зоны. В течение третьего триместра клетки этих тяжей продолжают расширяться и начинают продуцировать кортизол при регуляции адренокортикотропного гормона (АКТГ), определяющего возникновение пучкового слоя коры надпочечников взрослых.
Открытие, что альфа-рецептор эстрогена (ER-α ) и бета (ER-β ) находятся во всех зонах коркового вещества надпочечников плода позволяют предположить ,что эстрогены могут влиять непосредственно на функциональность надпочечников. Во время связывания с ER- α и ER- β эстрогены могут индуцировать воздействие на кору: прямое и косвенное ,и исходя из уровня эстрогенов в плазме крови они способны оказывать как стимулирующее, так и ингибирующее влияние на чувствительность клетки к АКТГ [20]. Было продемонстрировано, что при достижении высокого уровня эстрогенов в плазме, эстрогены способны оказывать действие на само корковое вещество надпочечников, для того, чтобы снизить его чувствительность к активности АКТГ. Это ингибирует продукцию ДГЭАС из области коры надпочечников плода и тем самым снижается синтез плацентарного эстрогена [20].
Сравнительные исследования полимеразной цепной реакции (ПЦР) в реальном времени показали, что экспрессия ER- α , ER- β и белка увеличивается с середины до поздней беременности в окончательной и переходной зонах надпочечников плода, но не в фетальной области [21]. Уменьшая количество доступных ER в области надпочечников плода, система отрицательной обратной связи, которая подавляет продукцию ДГЭАС, десенсибилизируется, активируется система опосредованной положительной обратной связи с эстрогенами, и продуцируется ДГЭАС.
Исследования, проводившиеся на долгосрочных культурах эмбриональных клеток надпочечников человека показали, что эстрогены связываются с ER- α и ER- β Могут оказывать стимулирующее действие синтез ДГЭАС с помощью косвенного пути, который снижает синтез кортизола [22]. Он является гормоном стресса, который оказывает поддерживающее воздействие на гомеостаз плода и влияет на созревание и развитие многих тканей, к которым относятся легкие, печень, кишечник и центральная нервная система [23]. При избыточном количестве кортизола в период беременности может возникать отрицательное влияние на развитие плода, при увеличении АД матери[23]. В качестве защитной реакции плода от избытка кортизола некоторые плацентарные ферменты 11 β –гидроксистероиддегидрогеназы имеют способность переводить его в неактивную стероидную форму-кортизон[20].Регуляцию этого плацентарного фермента осуществляют эстроген и прогестерон[24]. Повышение концентрации эстрогенов или прогестеронов стимулирует 11 β -гидроксистероиддегидрогеназы для увеличения трансплацентарного окисления кортизола до кортизона [24]. Пониженный уровень кортизола оказывает стимуляцию на секрецию АКТГ фетальным гипофизом, что стимулирующее влияет на выработку ДГЭАС в кортикальной части надпочечника плода, что обеспечивает основу для синтеза плацентарного эстрогена. В ситуациях, когда концентрация АКТГ становится высоким, петля обратной связи может быть отключена, что будет стимулировать синтез кортизола[20].
На более поздних этапах беременности экспрессия мРНК рецептора к АКТГ повышается в дефинитивной и переходной зонах, по отношению к эмбриональной зоне клеток надпочечников плода . Это свидетельствует о том, что АКТГ оказывает стимулирующее влияние на выработку кортизола в период поздней гестации. Синтез эстрогенов может ингибироваться посредством кортизола, при этом подавляется активность гипофизарного АКТГ. Но на поздних сроках беременности этот путь ,возможно ,не активируется, т.к. для инициации родов необходимо высокая концентрация эстрогенов. Вместо этого большое количество кортизола в плазме на поздних стадиях ,вероятно, посредством опосредованную эстрогенами систему положительной обратной связи преобразуются в кортизон.
По имеющимся данным удалось создать механистическое представление о влиянии эстрогенов на модуляцию в развитии и функционировании коркового вещества надпочечников[6,20]. Для того чтобы лучше понять влияние эстрогенов на надпочечниковую железу плода ,нужно охарактеризовать экспрессию коактиваторов и коэкспрессоров ,а так же структуру гетеродимеризации ER. Так же были идентифицированы три изоформы эстрогенов, однако точная роль их в физиологии беременности до сих пор не выяснена. Это может являться одним из ограничивающих факторов в современном понимании влияния эстрогенов на развитие надпочечников. Возможно структурные различия между 3 изоформами нужны для индуцирования селективных эффектов на уровне ядерных рецепторов. Последующие исследования в этой области необходимы для расшифровки роли эстрадиола ,эстрона и эстриола в функциональном и структурном развитии надпочечников плода.
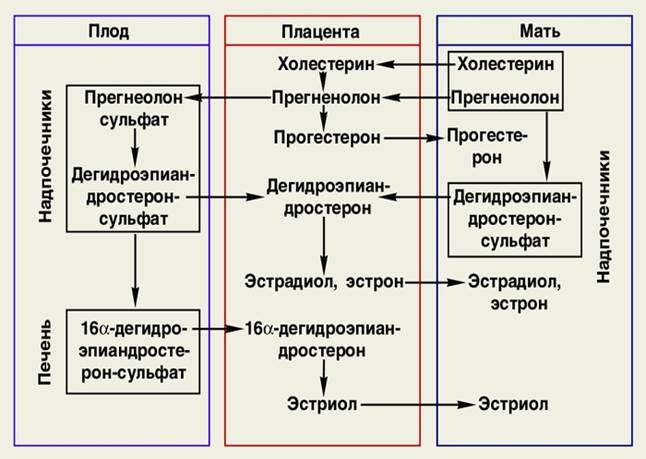
Рис 3:Синтез эстрогенов в плаценте и системе «мать-плод»
Лишь одно исследование занималось изучением влияния уровня эстрогена в плазме на дифференцировку коры[25]. В данной работе эмбриональные надпочечниковые железы были взяты у бабуинов с нормальным уровнем эстрогенов в плазме и у тех,кто имел аномально низкую концентрацию эстрогенов после введения высоко специфического ингибитора ароматазы. Эти исследования продемонстрировали ,что вес и объем надпочечников стали больше в 3 раза между серединой и концом беременности, что снизило уровень эстрадиола в сыворотке на 95%. 70%-ти процентный рост массы надпочечников плода среди бабуинов с низкими уровнями эстрогенов был объяснен значительным увеличением размеров плода за этот период беременности. Это интересный факт, потому что хотя ER- α и ER- β экспрессируются во всех зонах коры надпочечников плода [20], потеря эстрогена оказывает избирательное влияние на фетальную зону, первичный сайт продукции кортикостероидов. Наблюдаемое увеличение размера эмбриональной зоны было связано с 3-кратным повышением уровней ДГЭАС плода в сыворотке. Исходя из полученных результатов, исследователи пришли к выводу, что концентрация эстрогенов в плазме избирательно подавляет во втором и третьем триместре беременности как морфологическое, так и функциональное развитие матки [25]. Согласно этим выводам, возможно, что эстрогены, продуцируемый плацентой, возвращаются в кору надпочечников плода, чтобы сдержать ее рост и развитие. Оказывая контролирующее влияние на размер эмбриональной зоны, эстрогены способны регулировать количество выделяемого ДГЭАС, а так же собственное производство . Это имеет важное значение,т.к. высокий уровень эстрогена может приводить к нарушениям в работе эндокринной системы, что может оказывать пагубное влияние на рост и созревание плода [27].
Современные исследования показывают, что экзогенные эстрогены могут нарушать естественное взаимодействие между эстрогенами и корковым веществом надпочечников плода [25, 27]. Люди ежедневно подвергаются воздействию эстрогенов окружающей среды через продукты питания, в которых могут содержаться следы использующихся пестицидов, гербицидов, побочных продуктов переработки нефти и пластмасс, например, изофлавоны сои, бисфенол А, ДДТ, полихлорированные бифенилы, полибромированные дифениловые эфиры и др.[28]. Не смотря на то, что эстрогены представляют минимальную угрозу для взрослых, они так же могут оказывать отрицательное влияние на здоровье, если его воздействие приходится на критические стадии развития[4].
Воздействие эстрогенов окружающей среды во время перинатального развития может представлять собой постоянные долгосрочные последствия для всеобщего роста и развития[26]. У плода более высокая скорость метаболизма, более слабо развитый иммунитет и относительно низкая концентрация детоксицирующих ферментов и метаболитов печени ,если сравнивать со взрослым человеком, что обусловливает более высокую чувствительность к изменениям окружающей среды[29]. К тому же из-за небольшого размера тела плода влияние различных экзогенных эстрогенов может оказать пагубное влияние в виду химической интоксикации при концентрации, которая бы не оказала никакого неблагоприятного воздействия на взрослый организм[29]. В данный момент все больше [29]. Все большее число коммерчески доступных продуктов, содержащих низкие дозы эстрогенов, могут увеличивать риск накопления их в организме матери. Фетальные клетки являются очень подвержены воздействию ,поэтому влияние как эстрогенов окружающей среды, так и мощных химических веществ могут оказывать неблагоприятно воздействовать как на внутриутробный рост и развитие плода, так и на его будущее здоровье в целом.
Несмотря на то, что в наше время в окружающем нас мире много как экзогенных эстрогенов, так и эстрогеноподобных веществ, влияние, которое они оказывают на развитие плода, а так же на формирование надпочечниковых желез, активно не исследовалось. Исключением служит диэтилстильбэстрол (DES), синтетический эстроген. Его назначали беременным женщинам с середины 1940-х до 1970-х годов для того, чтобы предотвратить выкидыш. Использование данного препарата было прекращено после того, как было обследовано несколько миллионов потомков, и было обнаружено, что при внутриутробном воздействии DES вызывает серьезные изменения в развитии потомства, в первую очередь половой системы.
Потомство женского пола демонстрировало снижение фертильности и увеличение риска возникновения вагинальных карцином ,в то время как у потомства противоположного пола наблюдалось уменьшение размера яичек и снижение количества сперматозоидов[30].Плоды обоих полов, которые подвергались действию DES во время внутриутробного развития, оказались склоны к доброкачественным опухолям репродуктивных органов и к аутоиммунным заболеваниям[29]. Исходя из этого, изменения в развитии ,возникшие после внутриутробного воздействия DES, оказывают длительное влияние на программные функции репродуктивной системы. Как было написано раньше, клетки, являющиеся источником развития репродуктивных органов, являются теми же клетками, которые приводят к появлению эмбриональной надпочечной железы [6]. Предполагается, что пренатальное воздействие DES может также нарушать развитие надпочечников и их функции. У крыс, получавших DES с 8-го по 18-й день беременности, зарегистрировано увеличение массы надпочечников на 30% и при этом более низкая концентрация циркулирующих гормонов надпочечников по сравнению с контрольной группой, а также снижение концентрации прогестерона и эстрогена на 60 и 32%, соответственно. Было показано, что воздействие DES также уменьшает сократительную способность матки в родах, предотвращает отслойку плаценты от стенки матки и увеличивает смертность плодов.
При исследовании выживаемости плодов установлено, что плод крысы оказался наиболее чувствительным к лечению DES,если они воздействуют в период между 18 и 20 сутками беременности, и это указывает на то, что эстрогены окружающей среды оказывают наиболее сильное влияние на более поздних сроках беременности. Это вполне логично, т.к. DESпо своей структуре схож с эстрадиолом и может связываться с ER, чтобы индуцировать эстрогенподобные эффекты. Воздействие экзогенных эстрогенов на позднем сроке беременности может активировать цикл отрицательной обратной связи для ингибирования продуцирования ДГЭАС в коре надпочечников плода т.к. плод уже был подвержен влиянию высоких концентраций эндогенных эстрогенов . Более того, столь высокие уровни эстрогена могут способствовать снижению гонадотропинов в крови (к ним относятся ФСГ,ЛГ),и это в свою очередь может привести к торможению развития фолликулярных клеток и подавлять фолликулярное образование прогестерона и тестостерона . Фермент ароматаза способствует превращению тестостерона в эстрадиол ,и тем самым, этот путь может быть также отключен. Снижение функций фетальной коры и фолликулярного синтеза тестостерона в комбинации могут подавлять синтез de novo эстрогена. В настоящее время DES перестали использовать в клиническом аспекте для предотвращения выкидышей ,но он является очень полезным соединением ,которое позволяет исследовать роль и влияние эстрогенов на развитие плода. Результаты исследования DES сейчас используются для лучшей характеристики и понимания биологических эффектов иных эстрогенных соединений, в число которых так же входят пищевые эстрогены.
Заключение: Изучение уровня влияния эстрогенном эндогенной и экзогенной природы на правильное развитие и функционирование надпочечников необходимы для установления потенциальной роли женских половых гормонов в развитии эндокринных заболеваний, а также нарушений функционирования иммунной системы и регуляции метаболических процессов стероидными гормонами надпочечников.
Библиографическая ссылка
Шаповалова Е.Ю., Жабура М.О., Шостаковская А.С. ОСОБЕННОСТИ ПРЕНАТАЛЬНОГО ОНТОГЕНЕЗА НАДПОЧЕЧНИКОВ ПОД ВОЗДЕЙСТВИЕМ ЭКЗО- И ЭНДОГЕННЫХ ЭСТРОГЕНОВ // Международный студенческий научный вестник. – 2018. – № 6. ;URL: https://eduherald.ru/ru/article/view?id=19317 (дата обращения: 20.04.2024).